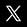|
|
|
|
 |
|
A D N: DU CALCUL Ã L'ASSEMBLAGE |
|
|
| |
|
| |
ADN : du calcul à l'assemblage
Bernard Yurke dans mensuel 347
daté novembre 2001 - Réservé aux abonnés du site
Depuis une dizaine d'années, on sait réaliser directement des calculs complexes en manipulant des fragments d'ADN. Mais l'apport principal de la biologie moléculaire à l'informatique résidera vraisemblablement dans l'assemblage automatique de circuits intégrés dont chaque composant ne sera pas plus gros qu'une molécule.
Si vous avez acheté un jour un ordinateur personnel, vous en avez fait l'expérience : la puissance de calcul de ces machines s'améliore bien plus vite que n'importe quelle autre caractéristique de n'importe quel autre produit grand public. Quelques années suffisent pour ravaler un appareil dernier cri au rang d'antiquité ! Cette progression n'est pas nouvelle. Depuis une quarantaine d'années que des ingénieurs fabriquent des circuits intégrés en silicium, le nombre de transistors par unité de surface ne cesse d'augmenter de façon exponentielle, doublant à peu près tous les deux ans.
Les dix prochaines années seront-elles aussi florissantes ? Beaucoup en doutent, car si la réduction de la taille des composants électroniques se poursuit au même rythme, ceux-ci ne seront bientôt constitués que de quelques atomes chacun. Saurons-nous surmonter toutes les difficultés qui se présenteront alors pour les fabriquer ? Des physiciens et des chimistes ont décidé d'attaquer le problème à sa base : plutôt que de réduire progressivement la taille des composants actuels, ils ont imaginé de se placer d'emblée à la plus petite échelle possible, et de construire des calculateurs directement avec des molécules.
A vrai dire, ils n'ont fait ainsi que copier des principes existants. L'évolution naturelle a en effet depuis quelques milliards d'années sélectionné des procédés de stockage et de manipulation des informations qui résolvent des problèmes aussi complexes que se nourrir, se déplacer, croître ou se reproduire. L'un des principaux supports de ces traitements d'informations chez les êtres vivants est l'ADN : dans nos gènes, c'est cette molécule qui contient les informations biologiques spécifiques à chacun d'entre nous. Pourrait-on utiliser l'ADN comme support d'un nouveau type d'informatique ?
Information naturelle. L'ADN a plusieurs qualités pour cela. D'abord, c'est un moyen de stockage d'informations compact. La molécule d'ADN ressemble en effet à une longue chaîne, un brin, dont l'un des côtés porte, à intervalles réguliers, des groupements chimiques que l'on nomme des bases, et dont il existe quatre modèles différents : l'adénine, la cytosine, la guanine et la thymine. Les informations génétiques sont codées par l'ordre dans lequel apparaissent ces bases. Chaque base mesure à peine un nanomètre de long, et les bases sont espacées d'un tiers de nanomètre environ. Si l'on imaginait par exemple de coder chaque caractère imprimé sur cette page par une suite de trois bases ce qui permet de coder 64 signes différents, on obtiendrait un brin d'ADN d'à peine vingt micromètres de long.
L'intérêt de l'ADN pour le calcul réside aussi dans la façon particulière avec laquelle les brins interagissent deux à deux. Si leurs séquences de bases sont complémentaires, c'est-à-dire si chaque adénine d'un brin correspond à une thymine de l'autre brin, et chaque cytosine à une guanine, alors les brins se lient l'un à l'autre et s'enroulent, formant une double hélice : on dit qu'ils s'hybrident c'est d'ailleurs sous cette forme de double hélice que l'on rencontre l'ADN la plupart du temps dans les organismes biologiques. Cette hybridation est spécifique : deux brins qui ne sont pas complémentaires ne peuvent pas se lier l'un à l'autre. Cette propriété permet de concevoir des brins d'ADN qui, lorsqu'on les mélange dans un tube à essai, s'assemblent en structures complexes prédéterminées.
Enfin, l'usage de l'ADN pour réaliser des calculs est favorisé par l'existence d'une « boîte à outils » qui permet de manipuler et de modifier les chaînes de bases. Les constituants de cette boîte à outils sont les enzymes, de grosses molécules, parmi lesquelles par exemple les ligases épissent deux brins d'ADN ensemble, les enzymes de restriction coupent un brin à un endroit précis, défini par une suite de bases spécifique, et les polymérases fabriquent des copies des brins. On dispose aussi, grâce aux développements de la biologie moléculaire, de nombreuses techniques de séparation et de purification de l'ADN.
En utilisant toutes ces qualités de l'ADN, Leonard Adleman, de l'université de Southern California a montré en 1994 que cette molécule permettait de résoudre un problème de calcul difficile, en l'occurrence le problème du chemin hamiltonien1. De quoi s'agit-il ? Imaginons une île avec un réseau de routes, dont certaines sont à sens unique, et dont quelques-unes en enjambent d'autres à l'aide de ponts. Les points de départ et d'arrivée étant donnés, existe-t-il un chemin qui passe une fois, et une fois seulement, par chaque croisement, tout en respectant les sens uniques ?
Calcul chimique. L. Adleman a codé les routes et les intersections avec des brins d'ADN, en particulier de façon que les brins représentant des routes qui se croisent aient à leurs extrémités des séquences complémentaires, qui leur permettent de s'hybrider. Lorsqu'il a mélangé ces brins, ils se sont assemblés en longues doubles hélices, dont chacune représentait un chemin respectant les sens uniques. Il obtenait ainsi tous les chemins possibles en une seule opération. Bien entendu, il fallait ensuite séparer toutes les doubles hélices, afin de repêcher celles qui correspondaient à un chemin hamiltonien, mais le nombre d'étapes nécessaires à cette opération n'augmente que de façon polynomiale avec le nombre d'intersections. Avec un ordinateur numérique, le nombre d'étapes nécessaires à la résolution de ce problème est beaucoup plus grand, et il augmente plus vite avec le nombre d'intersections.
Depuis, d'autres méthodes analogues ont été proposées pour résoudre avec de l'ADN divers problèmes complexes de calcul. Chaque fois, leurs promoteurs faisaient remarquer que le parallélisme massif, qui résulte de l'interaction simultanée d'un grand nombre de brins d'ADN différents, devrait permettre à un ordinateur à ADN de surpasser les ordinateurs électroniques.
Cette proposition est toutefois restée à l'état d'hypothèse. D'abord, les calculs effectivement réalisés avec de l'ADN ont porté sur des problèmes certes complexes, mais toujours dans des cas simples. L. Adleman, par exemple, n'a démontré expérimentalement la validité de sa méthode qu'en résolvant un problème de chemin hamiltonien à six intersections.
Lenteur rédhibitoire. Plusieurs difficultés pratiques ont aussi tempéré l'enthousiasme initial. D'abord, l'hybridation de l'ADN est un mécanisme infiniment lent par rapport à la fréquence des horloges des microprocesseurs actuels. En outre, plus le problème est complexe, plus le nombre de brins différents est grand : le volume d'ADN nécessaire croît souvent plus qu'exponentiellement avec la taille du problème à résoudre. Cela est dû au fait que, par exemple pour le problème du chemin hamiltonien, il faut construire effectivement tous les chemins possibles, dont le nombre augmente extrêmement vite avec le nombre d'intersections. Or, plus les brins d'ADN sont nombreux, plus ils mettent de temps à trouver un partenaire avec lequel ils puissent s'hybrider.
Je crains d'ailleurs que ce problème de vitesse ne soit un handicap fondamental vis-à-vis de l'ordinateur électronique. Quoi que l'on fasse, les électrons seront toujours plus mobiles, plus rapides, que des brins d'ADN, sauf à communiquer à ceux-ci une énergie considérable.
Enfin, les enzymes utilisées pour manipuler l'ADN font parfois des erreurs. Les méthodes de séparation et de purification ne sont pas parfaites non plus, et elles sont d'autant plus complexes que le nombre de brins à séparer est grand.
L'ordinateur à ADN ne serait-il alors qu'un gadget inutilisable, simple curiosité de laboratoire que les gens sérieux devraient s'empresser d'oublier ? Il est encore prématuré de conclure. Mais dans l'avenir proche, le principal intérêt du calcul avec l'ADN résidera à mon avis sur un autre terrain : l'assemblage d'ordinateurs électroniques à l'échelle moléculaire. Il n'est pas là en concurrence avec le calcul électronique : on ne peut pas construire d'objets physiques simplement en faisant circuler des électrons dans des plaques de silicium !
Cette idée a en fait été émise avant même que les premiers calculs ne soient réalisés avec de l'ADN. Dès 1987, Bruce Robinson, de l'université de Washington, et Nadrian Seeman avaient proposé d'assembler un réseau de mémoire à l'aide d'ADN2. Ils imaginaient que des blocs d'ADN pourraient s'assembler en réseaux cristallins tridimensionnels. Ces blocs porteraient les molécules analogues à des composants électroniques, fils électriques ou commutateurs par exemple, auxquelles les chimistes songeaient déjà à l'époque voir l'article de Christian Joachim, Jean-Pierre Launay et Ramon Companó. Leur proposition était toutefois restée théorique, faute de composants moléculaires adéquats, et faute de savoir concrètement comment assembler des blocs d'ADN.
C'est sur ce deuxième point que les résolutions de calculs avec de l'ADN ont permis de progresser. La relation entre le calcul et l'assemblage est en effet étroite. Dans chaque cas, on suit une procédure, un algorithme. Nous avons vu, avec l'exemple du chemin hamiltonien, que la mise au point d'une méthode de calcul par ADN est surtout la transformation d'un problème de mathématiques en un problème d'assemblage d'ADN. La spécificité de l'hybridation de l'ADN rend possible qu'un tel assemblage se déroule automatiquement dans le mélange. On pourrait donc utiliser les méthodes de calcul par ADN pour programmer l'auto-assemblage algorithmique de nanostructures.
Comment concrètement passer des calculateurs à ADN à l'assemblage de nanostructures ? L'idée originale de B. Robinson et de N. Seeman était d'utiliser des blocs tridimensionnels. Nous n'en sommes pas encore là, et nous nous contentons pour l'instant, ce qui n'est déjà pas si mal, d'apprendre à assembler des blocs bidimensionnels, des structures nommées tuiles d'ADN imaginées par Erik Winfree, de Caltech3.
Une tuile d'ADN est formée de deux doubles hélices entrecroisées : chaque brin d'une double hélice devient à un endroit membre de l'autre double hélice. A leurs extrémités, les deux doubles hélices se prolongent chacune par deux brins simples, qui ne sont pas complémentaires l'un de l'autre, et qui peuvent donc s'hybrider avec des brins portés par d'autres tuiles voir la figure ci-dessus. Pour le problème qui nous occupe, on peut se contenter de décrire ce type de structure de façon plus abstraite, comme une tuile d'où sortent quatre brins d'ADN, voire simplement comme une tuile dont chacun des bords est pourvu d'une attache colorée. Une tuile ne s'accroche à une autre que si les deux bords en contact portent des attaches de même couleur. Avec deux types de tuiles d'où émergent des brins complémentaires dont les attaches sont de même couleur, Erik Winfree et l'équipe de Nadrian Seeman, à l'université de New York, ont, les premiers, fait croître en 1998 des feuillets d'ADN qui ont la régularité d'un réseau cristallin simple4. Avec plus de tuiles et des règles d'interaction un peu plus complexes, on peut construire des structures plus élaborées. On y parvient en choisissant un ensemble approprié de séquences de bases pour les différents brins d'accrochage des tuiles. Les tuiles peuvent ainsi être programmées pour réaliser un calcul ou, ce qui revient au même, pour former un pavage prédéterminé voir l'encadré : « Des tuiles à calculer ».
L'industrie électronique pourrait-elle réellement utiliser cette technique pour assembler des cir- cuits ? Il faudrait pour cela maîtriser le problème de formation des défauts. E. Winfree a étudié ce problème d'un point de vue théorique5. La formation d'un pavage en tuiles d'ADN est analogue à la croissance d'un cristal. Or, la meilleure façon d'éviter la formation de défauts dans un cristal est de faire croître celui-ci à une température juste inférieure à son point de fusion, c'est-à-dire dans des conditions où le cristal, solide, est presque en équilibre thermodynamique avec le liquide. Autour d'un défaut, le cristal fond en effet à une température légèrement inférieure à son point de fusion : si la croissance est assez lente, les défauts qui se forment ont donc le temps de fondre et de disparaître au fur et à mesure.
Fusion d'ADN. Aux températures proches du point d'ébullition de l'eau, tous les brins d'ADN, même complémentaires, restent séparés les uns des autres. Puis, à mesure que la température diminue, les brins complémentaires d'ADN s'hybrident, et le restent de plus en plus longtemps. On peut définir le point de fusion d'une double hélice d'ADN comme la température où la moitié des brins complémentaires est hybridée les uns avec les autres. A cette température, les brins d'ADN s'associent aussi vite qu'ils se séparent. Cette température dépend de la concentration de l'ADN dans la solution, puisque plus le système est dilué, plus il faut de temps à deux brins complémentaires pour se rejoindre et s'hybrider.
De la même façon, pour un ensemble de tuiles tenues ensemble par des brins d'ADN complé- mentaires qui ont tous la même force de liaison, il existe une température de fusion, où la vitesse avec laquelle elles sont incorporées dans le réseau est égale à la vitesse à laquelle elles s'en séparent. En réalisant l'auto-assemblage à une tempéra- ture légèrement inférieure à cette température de fusion, on devrait obtenir des structures sans défauts.
La conséquence en est que moins on tolère de défauts dans la structure finale, plus il faut travailler près de la température de fusion et avec une faible concentration de tuiles, de façon que les défauts aient le temps de fondre. La croissance en est d'autant plus ralentie. Ce résultat n'est pas agréable pour les industriels, qui cherchent toujours à fabriquer leurs produits le plus vite possible pour une qualité donnée. Encore une fois, des limitations intrinsèques rendraient-elles l'ADN inutilisable pour l'électronique moléculaire ?
L'observation de ce qui se passe chez les êtres vivants nous laisse penser que non : la réplication de l'ADN dans les cellules, qui se déroule rapidement, dans des conditions bien différentes de l'équilibre thermodynamique, ne produit pourtant pas plus d'une erreur sur cent millions. Cette réplication de l'ADN est réalisée par une machine moléculaire nommée ADN polymérase qui, en plus de fabriquer la copie complémentaire d'un brin d'ADN, vérifie la conformité de cette copie avec l'original, et corrige les éventuelles erreurs.
Pourrions-nous fabriquer et utiliser des machines moléculaires aussi performantes pour l'assemblage algorithmique ? Nous sommes encore loin de savoir le faire. Mais nous pouvons essayer d'apprendre : c'est dans ce but que je me suis intéressé aux moteurs moléculaires en ADN. Mon équipe a mis au point des moteurs moléculaires simples, mus par l'énergie libérée lorsque deux brins d'ADN complémentaires s'hybrident et forment une double hélice6. Nous avons utilisé ces moteurs pour ouvrir et fermer une structure en V, une sorte de « pince moléculaire » faite aussi en ADN. L'ADN est un matériau très adaptable, qui peut être utilisé pour l'assemblage de structures complexes, mais aussi de structures complexes contenant des parties mobiles.
Dans tous les cas, l'auto-assemblage algorithmique ne remplacera pas du jour au lendemain la technologie de lithographie utilisée aujourd'hui pour la gravure des microprocesseurs en silicium. Cette dernière permet d'entasser cinquante millions de transistors sur un carré de 2 centimètres de côté, et ce chiffre n'a pas encore fini d'augmenter. La maîtrise acquise avec cette technologie en fait un concurrent redoutable pour la fabrication d'ordinateurs électroniques : nous ne sommes pas près de produire des structures aussi complexes que les circuits actuels, et avec une aussi bonne fiabilité, par auto-assemblage algorithmique. En outre, les composants moléculaires que l'on pourrait assembler de cette façon en les accrochant sur les tuiles, suivant la proposition initiale de B. Robinson et de N. Seeman, sont tout juste fabriqués dans les laboratoires de chimie.
Structures compactes. Toutefois, comme une double hélice d'ADN a un diamètre de 2 nanomètres, et que deux bases successives sont espacées de 0,34 nanomètre, les tuiles d'ADN ont réellement une taille nanométrique. Celles d'E. Winfree et N. Seeman ont des dimensions de 2 nanomètres par 4 et par 13 : c'est beaucoup plus petit que la largeur des transistors actuels, qui atteint environ 100 nanomètres. Comme les molécules utilisables comme composants ont une longueur de l'ordre du nanomètre, on peut envisager de placer dans des microprocesseurs moléculaires une densité de composants beaucoup plus grande que dans les microprocesseurs actuels.
Plus modestement, l'électronique pourrait entrer dans l'échelle moléculaire par une approche hybride, qui combinerait des techniques de lithographie actuelles et les techniques d'auto-assemblage moléculaire. De la même façon que les circuits intégrés sont, dans nos ordinateurs, connectés à des cartes qui sont, à leur tour, connectées à un châssis, on pourrait greffer de petits circuits électroniques moléculaires sur les circuits intégrés eux-mêmes. Cette technique permettrait dans un premier temps l'utilisation de circuits moléculaires simples, puis de plus en plus complexes à mesure que nous apprendrons à mieux manipuler l'ADN.
1 L.M. Adleman, Science , 266 , 1021, 1994.
2 B.H. Robinson et N.C. Seeman, Protein Engineering , 1 , 295, 1987.
3 E. Winfree, in DNA Based Computers, « Proceedings of a DIMACS Workshop », 4 avril, 1995, Princeton University, R.J. Lipton et E. B. Baum éd., volume 27 of DIMACS : Series in Discrete Mathematics and Theoretical Computer Science , p. 199-221, Providence, 1996. American Mathematical Society.
4 E. Winfree et al., Nature , 394 , 539, 1998.
5 E. Winfree, Ph. D. Thesis, California Institute of Technology 1998.
6 B. Yurke et al., Nature , 406 , 605, 2000.
7 C.T. Clelland, V. Risca, et C. Bancroft, Nature , 399 , 533, 1999.
8 P.K. Rothemund et E. Winfree, The Thirty-Second Annual ACM Symposium on Theory of Computing STOC 2000, 21-23 mai, Portland, Oregon USA, 2000.
DES TUILES À CALCULER
Comment programmer des tuiles d'ADN pour s'auto-assembler ? Eric Winfree, de Caltech, et Paul Rothemund, de l'université Southern California8, ont imaginé un procédé pour écrire en binaire la liste des nombres jusqu'à une valeur prédéterminée en utilisant de telles structures. Ainsi 24 tuiles sont nécessaires pour compter jusqu'à 15 A. Les règles d'assemblage sont les suivantes : une tuile ne peut adhérer à la structure en construction par un bord seulement que si la couleur rouge y est présente, et que les autres couleurs correspondent aussi ; s'il n'y a pas de rouge sur les bords de la structure, une tuile ne peut être ajoutée que si elle a deux bords en contact, et que les couleurs correspondent sur ces deux bords ; si un bord est complètement vert, il ne peut coller à aucun autre bord ; si un bord porte un chiffre, la tuile n'adhère à la structure que si les chiffres et les couleurs correspondent tous les deux.
On fabrique des tuiles d'ADN qui obéissent à ces règles de construction en choisissant convenablement les séquences de base et la longueur des brins, de façon que les brins correspondant aux bords de tuile en rouge se lient plus fortement que les autres. On y parvient car la liaison entre la guanine et la cytosine est plus forte que celle entre l'adénine et la thymine, et parce que des brins d'ADN complémentaires se lient d'autant plus fortement qu'ils sont longs. Lorsque l'on a deux séries de brins avec deux forces de liaison différentes, on peut trouver une température à laquelle les brins les plus faiblement liés ne maintiennent une tuile en place que si les brins de deux autres bords ou plus participent à la liaison. A cette température, l'auto-assemblage se déroule conformément aux règles. Les tuiles qui s'assemblent pour former la première ligne ont du rouge sur leurs bords de contact. Lorsque la première structure dessinée est assemblée, deux bords de contact deviennent disponibles, et la rangée suivante est complétée. Ainsi, l'assemblage procède par allers et retours entre deux bords B. Les 24 tuiles différentes de l'ensemble ont été choisies de façon qu'à un certain point aucune autre tuile ne peut plus être ajoutée parce qu'il n'y a plus de bords correspondants : la croissance s'arrête.
Le motif bleu foncé du pavage d'ADN correspond à la disposition des 1 dans la suite des nombres de 0 à 15 écrits en binaire C. Le choix judicieux des couleurs sur les différents bords des tuiles a donc permis de les programmer pour écrire une liste de nombres en binaire de 0 à 15. En augmentant un peu le nombre de types de tuiles, en particulier en ajoutant des tuiles avec des paires de nombres 0,1, 1,2, 2,3, 3,4, ... n-1, n, les tuiles écriront de 0 à 2n - 1.
L'assemblage de pavages analogues permet d'exécuter des opérations mathématiques et logiques. L'avantage de l'auto-assemblage algorithmique est que le nombre de types de tuiles est souvent bien inférieur au nombre total de tuiles nécessaire. Par exemple, pour le compteur binaire présenté ici, 20 + n types de tuiles sont utilisés pour fabriquer un pavage de n par 2n + 1+ 1 tuiles.
Ce pavage d'ADN peut être utilisé comme support d'un circuit électronique, en accrochant sur les tuiles les composants adéquats. Si l'on indique en entrée en bas du pavage un nombre binaire correspondant à l'une des lignes, la valeur de sortie de cette ligne, et de cette ligne seulement, passe de 0 à 1, ce qui permet de commander une cellule mémoire.
SAVOIR
A lire :
-N . C. Seeman , Acc. Chem. Res., 30 , 357, 1997.
-C.M. Niemeyer, Angew. Chem. Int. Engl. , 36 , 585, 1997.
-C.M. Niemeyer, Appl. Phys. A , 68 , 119, 1999.
DOCUMENT la recherche.fr LIEN |
| |
|
| |
|
|
|
CELLULES GLIALES |
|
|
| |
|
| |
Cellule gliale
Dans le système nerveux, les cellules gliales (parfois nevroglie ou tout simplement glie, du grec γλοιός (gloios), « gluant ») sont les cellules qui forment l'environnement des neurones. Elles assurent le maintien de l'homéostasie, produisent la myéline et jouent un rôle de soutien et de protection du tissu nerveux en apportant les nutriments et l'oxygène, en éliminant les cellules mortes et en combattant les pathogènes.
Les cellules gliales représentent environ 50 % du volume cérébral1 et au plus 50 % des cellules du cerveau2, contrairement à l'assertion très répandue affirmant des ratios de 10:1 à 50:13, sans aucune référence sérieuse. On distingue en général 4 principaux types de cellules gliales :
• les astrocytes ;
• les oligodendrocytes ;
• les cellules de Schwann ;
• la microglie.
Contrairement à la grande majorité des neurones, les cellules gliales peuvent se diviser par mitose.
Pendant longtemps, l'implication des cellules gliales dans le traitement de l'information nerveuse a été ignorée par rapport au rôle proéminent des neurones, mais il est aujourd'hui reconnu qu'elles exercent une action modulatrice sur la neurotransmission bien que le détail de ces mécanismes reste mal compris.
Astrocyte
Les astrocytes sont des cellules gliales du système nerveux central. Elles ont généralement une forme étoilée, d'où provient leur étymologie : Astro - étoile et cyte - cellule. Elles assurent une diversité de fonctions importantes, centrée sur le support et la protection des neurones. Ces cellules participent au maintien de la barrière hémato-encéphalique, régulent le flux sanguin, assurent l'approvisionnement en nutriments et le métabolisme énergétique du système nerveux, participent à la neurotransmission et maintiennent la balance ionique du milieu extracellulaire. Les astrocytes jouent également un rôle dans la défense immunitaire, la réparation et la cicatrisation du cerveau ou de la moelle épinière après une lésion3.
Les recherches récentes révèlent la complexité et l'importance de cette population cellulaire. Les astrocytes sont capables d'assurer une certaine forme de communication, reposant sur des vagues intracellulaires de Ca2+, et peuvent également libérer certains neurotransmetteurs (appelés gliotransmetteurs). Cette forme de communication, que l'on croyait spécifique aux neurones, leur confèrerait un rôle beaucoup plus actif dans le fonctionnement du cerveau, notamment sur la plasticité des communications neuronales. Il a également été montré que certaines sous-populations d'astrocytes ont des propriétés de cellules souches neurales et sont à la source du mécanisme de neurogenèse adulte.
Oligodendrocyte
Un oligodendrocyte est une cellule de la névroglie interstitielle. Sa principale fonction est la formation de la gaine de myéline entourant les fibres nerveuses (axones) du système nerveux central (SNC); la formation de la myéline au niveau du système nerveux périphérique étant assurée par les cellules de Schwann. La gaine de myéline permet d'augmenter la vitesse de propagation et la fréquence des influx nerveux. Un seul oligodendrocyte est capable de myéliniser jusqu'à 50 axones1.
Cellule de Schwann
Les cellules de Schwann (ou neurolemmocytes) sont une variété de cellules gliales qui assurent principalement l'isolation myélinique des axones du système nerveux périphérique des vertébrés (on les classe donc parmi les « cellules gliales périphériques »). Comme les oligodendrocytes du système nerveux central, elles assurent la myélinisation—c'est-à-dire l'isolation électrique—des axones mais dans le système nerveux périphérique. Il existe néanmoins de petites différences entre ces deux types de cellules.
Leur nom est un hommage au physiologiste allemand Theodor Schwann.
Microglie
La microglie (ou microgliocytes) est une population de cellules gliales constituée de macrophages résidents du cerveau et de la moelle épinière formant ainsi la principale défense immunitaire active du système nerveux central.
Cellule gliale
Dans le système nerveux, les cellules gliales (parfois nevroglie ou tout simplement glie, du grec γλοιός (gloios), « gluant ») sont les cellules qui forment l'environnement des neurones. Elles assurent le maintien de l'homéostasie, produisent la myéline et jouent un rôle de soutien et de protection du tissu nerveux en apportant les nutriments et l'oxygène, en éliminant les cellules mortes et en combattant les pathogènes.
Les cellules gliales représentent environ 50 % du volume cérébral1 et au plus 50 % des cellules du cerveau2, contrairement à l'assertion très répandue affirmant des ratios de 10:1 à 50:13, sans aucune référence sérieuse. On distingue en général 4 principaux types de cellules gliales :
• les astrocytes ;
• les oligodendrocytes ;
• les cellules de Schwann ;
• la microglie.
Contrairement à la grande majorité des neurones, les cellules gliales peuvent se diviser par mitose.
Pendant longtemps, l'implication des cellules gliales dans le traitement de l'information nerveuse a été ignorée par rapport au rôle proéminent des neurones, mais il est aujourd'hui reconnu qu'elles exercent une action modulatrice sur la neurotransmission bien que le détail de ces mécanismes reste mal compris.
Astrocyte
Les astrocytes sont des cellules gliales du système nerveux central. Elles ont généralement une forme étoilée, d'où provient leur étymologie : Astro - étoile et cyte - cellule. Elles assurent une diversité de fonctions importantes, centrée sur le support et la protection des neurones. Ces cellules participent au maintien de la barrière hémato-encéphalique, régulent le flux sanguin, assurent l'approvisionnement en nutriments et le métabolisme énergétique du système nerveux, participent à la neurotransmission et maintiennent la balance ionique du milieu extracellulaire. Les astrocytes jouent également un rôle dans la défense immunitaire, la réparation et la cicatrisation du cerveau ou de la moelle épinière après une lésion3.
Les recherches récentes révèlent la complexité et l'importance de cette population cellulaire. Les astrocytes sont capables d'assurer une certaine forme de communication, reposant sur des vagues intracellulaires de Ca2+, et peuvent également libérer certains neurotransmetteurs (appelés gliotransmetteurs). Cette forme de communication, que l'on croyait spécifique aux neurones, leur confèrerait un rôle beaucoup plus actif dans le fonctionnement du cerveau, notamment sur la plasticité des communications neuronales. Il a également été montré que certaines sous-populations d'astrocytes ont des propriétés de cellules souches neurales et sont à la source du mécanisme de neurogenèse adulte.
Oligodendrocyte
Un oligodendrocyte est une cellule de la névroglie interstitielle. Sa principale fonction est la formation de la gaine de myéline entourant les fibres nerveuses (axones) du système nerveux central (SNC); la formation de la myéline au niveau du système nerveux périphérique étant assurée par les cellules de Schwann. La gaine de myéline permet d'augmenter la vitesse de propagation et la fréquence des influx nerveux. Un seul oligodendrocyte est capable de myéliniser jusqu'à 50 axones1.
Cellule de Schwann
Les cellules de Schwann (ou neurolemmocytes) sont une variété de cellules gliales qui assurent principalement l'isolation myélinique des axones du système nerveux périphérique des vertébrés (on les classe donc parmi les « cellules gliales périphériques »). Comme les oligodendrocytes du système nerveux central, elles assurent la myélinisation—c'est-à-dire l'isolation électrique—des axones mais dans le système nerveux périphérique. Il existe néanmoins de petites différences entre ces deux types de cellules.
Leur nom est un hommage au physiologiste allemand Theodor Schwann.
Microglie
La microglie (ou microgliocytes) est une population de cellules gliales constituée de macrophages résidents du cerveau et de la moelle épinière formant ainsi la principale défense immunitaire active du système nerveux central.
|
| |
|
| |
|
|
|
CERVEAU |
|
|
| |
|
| |
( consulter le fichier PDF en bas de la page )
1
AIRES FONCTIONNELLES
Sommaire :
LOBES et AIRES
LOBE FRONTAL : aires motrices
LOBE TEMPORAL : aires auditives, aires olfactives
LOBE OCCIPITAL : aires visuelles
LOBE PARIÉTAL : aires tactiles, aires gustatives FONCTIONS COGNITIVES – COGNITION
FONCTIONS LANGAGIÈRES – LANGAGE
FONCTIONS ÉMOTIONNELLES – ÉMOTIONS
FONCTIONS MNÉSIQUES – MÉMOIRES
******
Termes
- mnésie : fait référence à la mémoire
- amnésie : perte de la mémoire - gnosie : perception, capacité visuelle, auditive, olfactive, gustative, tactile à reconnaitre ou à identifier - sensoriel : renvoie aux cinq sens, les récepteurs sont regroupés dans un organe (vision, audition, olfaction et gustation).
- sensitif (sensibilité ou somesthésie) : renvoi à toutes les sensations du corps, les récepteurs ont une répartition plus ubiquitaire, ils sont distribués dans l’ensemble du corps.
Lorsque les experts font de l’imagerie IRM, on observe l’activation localisée dans tel lobe.
Il est possible de diviser sommairement et schématiquement le cortex cérébral en :
- aires primaires (aire motrice, aire sensorielle, aire sensitive)
- aires associatives (appelées aussi aires secondaires)
Les lobes sont divisés en aires fonctionnelles :
- aires motrices : motricité
- aires sensitives : somesthésie (tactile….)
- aires sensorielles : auditive, olfactive, visuelle, tactile, gustative
Localisation :
- aires motrices : lobe frontal
- aires sensitives : lobe pariétal
- aires sensorielles : lobe pariétal, temporal, occipital
Fonction :
- l’aire primaire traite les informations sensorielles et motrices brutes
- l’aire associative joue un rôle dans l'intégration de ces informations
Communication :
Presque chaque aire motrice, sensorielle, sensitive a une aire primaire et associative.
Les aires associatives reçoivent des afférences des aires primaires.
Les aires associatives interprètent les informations des aires primaires.
Les aires associatives entourent l’aire primaire correspondante (elles communiquent entre elles).
2
L’aire primaire est soit le point de départ, soit d’arrivée d’une fonction.
Il n’y a aucune fonction d’intégration.
L’aire secondaire ou associative est une aire d’intégration.
Il y a une fonction d’intégration.
Exemple :
Au niveau du lobe occipital, la fonction est la vision :
- l’aire visuelle primaire : permet de voir
- l’aire visuelle associative : permet de comprendre ce qu’on a vu
- les aires motrices sont des aires effectrices qui donnent les liaisons efférentes (l’information part du cerveau)
- les aires sensorielles sont des aires réceptrices qui ont des liaisons afférentes (l’information arrive au cerveau)
- les aires associatives envoient des commandes motrices aux muscles et aux glandes
Une aire associative est toujours située à proximité de l'aire primaire qui lui correspond.
Les aires associatives communiquent avec les aires sensorielles primaires et les aires motrices primaires. Les aires d’association jouent un rôle dans l'intégration de fonctions complexes telles que la mémoire, les émotions, la volonté, le jugement, l'intelligence ou la personnalité.
Le premier degré de traitement de l'information se situe près des aires sensorielles primaires.
Ainsi, les aires d'association visuelle, auditive, somesthésique interprètent les sensations reçues en fonction des apprentissages antérieurs (mémoire visuelle, auditive et somesthésique) et leur donnent du sens.
Le cortex associatif est aussi appelé aires associatives, car il est composé de différentes aires, auxquelles on attribue des fonctions différentes.
Les aires associatives se distinguent des autres aires car elles reçoivent des afférences de plus d’un système sensoriel (par exemple vision et audition…).
Les aires associatives participent donc à la genèse de notre perception du monde, qui intègre sans les décomposer les différentes modalités perceptives.
Le cortex associatif permet la formation de nos perceptions qui sont une interprétation de nos sensations, une mise en relation de ces sensations avec notre vécu, nos attentes et nos connaissances.
Le cortex préfrontal est la plus complexe des aires associatives et la dernière à se développer.
Il est relié à l'intelligence, à la cognition, au raisonnement et au jugement.
C'est lui qui permet à l'individu de s'adapter aux codes de la société dans laquelle il vit et de surseoir à la satisfaction du besoin immédiat.
Des lésions vasculaires ou tumorales dans cette région, ou sa désinhibition par l'ingestion d'alcool entraînent des troubles de la personnalité, des comportements inadaptés, voire des troubles psychiatriques.
Le cortex préfrontal est présent à toutes les étapes de la mémorisation (encodage, consolidation, rappel). Son rôle est à la fois d'orienter l'attention visuelle, de créer des liens entre les éléments mémorisés (donc apprendre) et l'inhibition des informations distrayantes.
Lorsque le cortex préfrontal est atteint, les capacités d'apprentissage et de mémorisation sont fortement perturbées (limites dans la quantité de mémoire, difficultés à faire des liens, enregistrement d'informations non pertinentes, etc.), comme c'est le cas dans les troubles de l'attention et l'hyperactivité.
3
LOBES CÉRÉBRAUX et AIRES CÉRÉBRALES
Le cortex cérébral est divisé en quatre lobes : frontal, temporal, occipital, pariétal
Chaque hémisphère à un lobe frontal, un lobe temporal, un lobe occipital, et un lobe pariétal.
Les fonctions du cerveau sont réparties entre ces lobes. Chaque lobe peut remplir plusieurs fonctions.
Une atteinte de l’une des zones entraine un dérèglement dans la fonction qui y est associée.
4 lobes externes : lobe frontal, lobe temporal, lobe occipital, lobe pariétal.
2 lobes cachés dans les replis corticaux : lobe limbique et lobe insulaire.
LOBES
AIRES
FONCTIONS
Frontal
aires motrices
cognition, motricité
Temporal
aires auditives, aires olfactives
ouïe, odorat
Occipital
aires visuelles
vue
Pariétal
aires gustatives, aire tactiles
goût, toucher, somesthésie
4
LOBE FRONTAL
Fonctions : la cognition
- Aires motrices volontaires
- Aires préfrontales
- Mémoire de travail
- Mémoire épisodique
- Mémoire sémantique
- Langage (expression) : Aire de Broca
- Ecriture
Précision
Motricité : il contient les centres chargés du contrôle musculaire, mais aussi des mouvements rythmiques coordonnés de la tête et de la gorge, comme ceux consistant à mâcher, lécher ou avaler.
LOBE TEMPORAL
Fonctions : l’audition, l’olfaction
- Aires auditives
- Aires olfactives
- Aire vestibulaire
- Mémoire épisodique
- Mémoire sémantique
- Langage (compréhension) : Aire de Wernicke
- Lecture
Précision
L’aire olfactive se trouve dans le système limbique localisé dans le lobe temporal.
LOBE OCCIPITAL
Fonctions : la vision
- Aires visuelles
- Mémoire épisodique
LOBE PARIÉTAL
Fonctions : le toucher et la sensibilité
- Aires tactiles
- Aires gustatives
- Aires sensitives
- Mémoire épisodique
- Habileté visuo-spatiale
Précision
L’aire sensitive = aire somesthésique = aire somatosensorielle
5
FONCTIONS COGNITIVES – COGNITION Cognition : tout ce qui concerne le traitement de l’information par le cerveau (y compris les émotions).
Les fonctions cognitives représentent tous les processus cérébraux/mentaux par lesquels on acquiert, traite, conserve, récupère, utilise l’information pour agir. Les principales fonctions cognitives sont : - le langage - les mémoires - les émotions - l'attention sous ses différentes formes - les gnosies : perception, reconnaissance, identification / capacité à savoir reconnaitre - les praxies : gestes / capacité à savoir faire - les habiletés visuo-spatiales et temporo-spatiales - les fonctions exécutives Précision Les habiletés visuo-spatiales permettent de percevoir les objets de notre environnement et de s’orienter dans l’espace.
Les habiletés temporo-spatiales permettent de se repérer/s’orienter par rapport au temps et à l’espace.
Les fonctions exécutives entrent en jeu dans : - l'inhibition (capacité à juger des informations non pertinentes pour la réalisation de la tâche) - le raisonnement - la logique - la pensée abstraite - la prise de décision - le jugement - l'anticipation - la planification - l'organisation - la motivation - l'initiative - la flexibilité - la résolution de problème - l'apprentissage de règles - la sélection de réponses motrices N.B. Pour préparer un repas, une réunion, un voyage, on se sert des fonctions exécutives (ex : planification…) et on a aussi besoin d’autres compétences cognitives (ex : mémoire, langage, habiletés visuo-spatiales…).
Les fonctions exécutives sont des fonctions cognitives.
Certaines fonctions cognitives ne sont pas des fonctions exécutives.
Mais les fonctions exécutives font partie des fonctions cognitives.
6
FONCTIONS LANGAGIÈRES – LANGAGE
Localisation du langage dans le cerveau
Paul Broca et Carl Wernicke (deux neurologues) ont découvert les aires du langage du cerveau.
- Aire de Broca (lobe frontal) : la zone de production des mots
- Aire de Wernicke (lobe temporal) : la zone de la compréhension (décodage) de ces mots (oraux ou écrits)
FONCTIONS ÉMOTIONNELLES – ÉMOTIONS
Localisation des émotions dans le cerveau
Le système limbique, qui comprend l’amygdale et plusieurs régions étroitement reliées, est principalement responsable du traitement des émotions.
FONCTIONS MNÉSIQUES – MÉMOIRES
Localisation des mémoires dans le cerveau
- Mémoire de travail liée au lobe frontal.
- Mémoire épisodique liée aux lobes frontal, temporal, occipital, pariétal, ainsi qu’à l’hippocampe et à l’amygdale.
- Mémoire sémantique liée aux lobes frontal, temporal.
- Mémoire procédurale liée au cervelet.
La mémoire n'est pas localisée dans un endroit précis dans le cerveau.
Il n'est pas possible de préciser l'endroit exact où se situe la mémoire dans le cerveau.
Les mémoires sont liées à différentes zones du cerveau.
******
7
SCHÉMAS
1 2 3 4 5 6 7
aire préfrontale aire motrice d'association aire motrice primaire aire somesthésique primaire aire somesthésique d'association aire visuelle d'association aire visuelle primaire
8
Lobe frontal
Lobe Temporal
9
Lobe Occipital
Lobe Pariétal
10
Aire motrice primaire (lobe frontal) : permet d’envoyer les ordres de la motricité volontaires aux muscles
Aire somesthésique primaire (lobe pariétal) : reçoit les sensations en provenance de la périphérie du corps
11
Cervelet
Corps calleux
12
Substance
Aire de Broca (lobe frontal) / Aire de Wernicke (lobe temporal)
FIN
DOCUMENT www.noesis-reseau.com LIEN |
| |
|
|
|
| |
|
|
|
SCHIZOPHRÃNIE |
|
|
| |
|
| |
CERVEAU ET PSY
Schizophrénie : la nicotine, un atout pour le cerveau ?
Le 26.01.2017 à 17h28
Courante chez les personnes atteintes de schizophrénie, l'addiction à la nicotine permettrait de compenser le déficit de fonctionnement de certaines cellules dans le cerveau.
La nicotine, un atout pour le cerveau des schizophrènes ?
Une large proportion des personnes atteintes de schizophrénie présente une forte dépendance à la nicotine. Pour nombre des 600.000 malades que compte la France, il s’agirait en fait d’une quasi auto-médication, cette substance pouvant améliorer l'activité des cellules du cortex préfrontal impliquées dans cette maladie psychiatrique. C'est ce que suggère une étude parue le 23 janvier 2017 dans Nature Medicine, réalisée notamment par des chercheurs français de l'Institut Pasteur, de l'Inserm et du CNRS. Avec d'autres scientifiques allemands, russes et américains, ils sont parvenus à décrypter, chez des souris modèles de la schizophrénie, le mode d'action de cette molécule psychostimulante sur les neurones du cortex préfrontal. Cette région située à l'avant du cerveau est le siège privilégié de différentes fonctions cognitives dites "supérieures" : raisonnement, langage, mémoire immédiate, etc.
Perte de contact avec la réalité, hallucinations auditives, paranoïa, bouffées délirantes... Ces symptômes sont les plus connus de la schizophrénie, et les plus impressionnants. Mais cette pathologie psychiatrique s'accompagne aussi de signes moins visibles, pourtant très invalidants et moins bien pris en charge : fonctions cognitives émoussées, perte d’attention, problèmes de mémoire de travail...
La nicotine rétablit l'activité des interneurones
Il y a peu, une mutation génétique, CHRNA5, codant pour une partie du récepteur nicotinique a été associée à la fois aux troubles cognitifs propres aux schizophrènes et à la dépendance au tabac. En introduisant ce gène humain dans des souris, les chercheurs ont reproduit ces déficits cérébraux caractéristiques de la maladie. Mieux, ils ont pu identifier le type de cellules du cortex préfrontal dont l'activité était diminuée par cette mutation : les interneurones, sorte de petits neurones qui établissent des connexions entre différents réseaux de neurones. Surtout, l'administration de nicotine chez ces modèles a montré une amélioration de l'activité de ces interneurones. C'est ce que les scientifiques ont pu constater in vivo grâce à une technique d'imagerie (voir vidéo ci-dessous).
« Les travaux portant sur ce modèle de la maladie montrent que lorsque nous administrons de la nicotine, celle-ci se fixe sur les récepteurs nicotiniques des interneurones, et influence l’activité des cellules pyramidales du cortex préfrontal qui retrouvent un état d’excitation normal », explique le Dr Fani Koukouli, première auteure de l’étude et chercheuse à l'Institut Pasteur. « Pour faire simple, la nicotine compense un défaut génétique déterminé. Personne n’a jamais montré cela avant », précise le Dr Jerry Stitzel, neuroscientifique à l'université du Colorado à Boulder (Etats-Unis).
Activité neuronale dans le cortex préfrontal de contrôle, à gauche ; et dans un cortex exprimant la mutation humaine CHRNA5, à droite © Institut Pasteur.
Les effets bénéfiques de cette administration répétée de nicotine laissent penser qu'une nouvelle cible thérapeutique contre la schizophrénie pourrait être développée. "La molécule thérapeutique devra alors présenter la même forme que la nicotine sans en avoir les effets nocifs (dépendance, vieillissement cellulaire, accélération de l’activité cardio-vasculaire…)", conclut Uwe Maskos, principal auteur de l’étude et responsable de laboratoire à l'Institut Pasteur.
DOCUMENT sciencesetavenir.fr LIEN |
| |
|
| |
|
| Page : [ 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 ] Précédente - Suivante |
|
|
|
|
|
|